Via het Linkedin account van Mario Ortiz Martinez vond ik de volgende productie waarin wordt uitgelegd dat het Pfizer vaccin het DNA van de lever aantast. Voorts wordt de Neurenberg Code aangehaald, waarin duidelijk is weergegeven dat het iedereen vrij moet staan om wel dan niet te kiezen voor een vaccinatie en gezien de tekst zit daar geen woord Spaans bij (overigens heeft de EU eerder laten weten tegen verpolichte vaccinatie te zijn, al is die mening intussen losgelaten door hare kwaadaardigheid von der Leyen, althans als ik me niet vergis):
“1. The voluntary consent of the human subject is absolutely essential. This means that the person involved should have legal capacity to give consent; should be so situated as to be able to exercise free power of choice, without the intervention of any element of force, fraud, deceit, duress, over-reaching, or other ulterior form of constraint or coercion.”
Deze tekst kom je hieronder nogmaals tegen en gezien de volledige tekst is het m.i. onverantwoord om je te laten vaccineren met het Pfizer vaccin. Maar lees de tekst en vorm je eigen oordeel. (zie alvorens tot vaccinatie over te gaan ook de berichten onder de door mij toegevoegde links en ook daarbij: vorm je eigen mening)
(On
the top right hand side of this page you can choose for a translation
in the language of your choice: choose ‘Engels’
[english] so you can recognise your own language [the Google
translation is first in dutch, a language most people don’t
understand, while on the other hand most people recognise there
language translated in english])
(als
je het Engels niet machtig bent, kopieer dan de Engelse tekst en plak
die in deze
vertaalapp,
de app werkt snel en de vertaling is van een redelijk goede
kwaliteit. Kopieer
het artikel en plaats het links in de app, je ziet rechts Italiaans
staan als je daar op klikt kan je kiezen voor Nederlands)
Farma Medical-Marketing Projects, Medical & Farma 3D animation videos, Director Herr Zimmerman – Events & Music.

Expert in Business Succession/Exiting Execution – Speaker – Author of The Business Novel – The King of Main Street
URGENT – BE IN FORMED – Please Read and Then Share
with Loved Ones: Study shows Pfizer COVID shot changes the liver cells
DNA according to Swedish researchers at Lund University – one of
Europe’s oldest and most prestigious research universities. These
findings are published in the current -March 2022- Issue of the
peer-reviewed journal – Molecular Biology.
***This first source full peer-reviewed article is included with this post below.***
ARTICLE
ABSTRACT: These researchers found that messenger RNA (mRNA)
from Pfizer’s COVID-19 gene therapy shot is able to enter
human liver cells and convert into DNA- combining with the human genome –
creating something – NEW – NEVER SEEN BEFORE!!!!!!
THIS IS CAUSE FOR ALARM – ARE HUMANS PATENABLE?
On
June 13, 2013 – the US Supreme Court ruled that DNA manipulated by
humans are eligible to be patented. This synthetic DNA is produced from
messenger RNA that serves as the instructions for making proteins in the
cell. Does this sound familiar? (i.e. – mRNA shots) – You can’t write
this stuff up.
Again, please read and then share this
document/article with as many people you care about as possible. Below
in the comments a link to a down loadable pdf has been provided.
FOOD FOR THOUGHT:
WHAT
IS INFORMED CONSENT – Point One of The Nuremberg Code – this is taken
directly from United States National Institutes of Health:
“1.
The voluntary consent of the human subject is absolutely essential. This
means that the person involved should have legal capacity to give
consent; should be so situated as to be able to exercise free power of
choice, without the intervention of any element of force, fraud, deceit,
duress, over-reaching, or other ulterior form of constraint or
coercion.”
Hier de productie van MDPI:
Intracellular Reverse Transcription of Pfizer BioNTech COVID-19 mRNA Vaccine BNT162b2 In Vitro in Human Liver Cell Line

1 ,
,
1,
1,
1,
1,
2 and
1,*
1
Department of Clinical Sciences, Lund University, 20502 Malmö, Sweden
2
Infection Medicine, Department of Clinical Sciences, Lund University, 22362 Lund, Sweden
*
Author to whom correspondence should be addressed.
Academic Editor: Stephen Malnick
Curr. Issues Mol. Biol. 2022, 44(3), 1115-1126; https://doi.org/10.3390/cimb44030073
Received: 18 January 2022
/
Revised: 19 February 2022
/
Accepted: 23 February 2022
/
Published: 25 February 2022
(This article belongs to the Topic Clinical, Translational and Basic Research on Liver Diseases)
Abstract
Preclinical studies of COVID-19 mRNA vaccine BNT162b2, developed by
Pfizer and BioNTech, showed reversible hepatic effects in animals that
received the BNT162b2 injection. Furthermore, a recent study showed that
SARS-CoV-2 RNA can be reverse-transcribed and integrated into the
genome of human cells. In this study, we investigated the effect of
BNT162b2 on the human liver cell line Huh7 in vitro. Huh7 cells were
exposed to BNT162b2, and quantitative PCR was performed on RNA extracted
from the cells. We detected high levels of BNT162b2 in Huh7 cells and
changes in gene expression of long interspersed nuclear element-1
(LINE-1), which is an endogenous reverse transcriptase.
Immunohistochemistry using antibody binding to LINE-1 open reading
frame-1 RNA-binding protein (ORFp1) on Huh7 cells treated with BNT162b2
indicated increased nucleus distribution of LINE-1. PCR on genomic DNA
of Huh7 cells exposed to BNT162b2 amplified the DNA sequence unique to
BNT162b2. Our results indicate a fast up-take of BNT162b2 into human
liver cell line Huh7, leading to changes in LINE-1 expression and
distribution. We also show that BNT162b2 mRNA is reverse transcribed
intracellularly into DNA in as fast as 6 h upon BNT162b2 exposure.
Keywords:
COVID-19 mRNA vaccine; BNT162b2; liver; reverse transcription; LINE-1; Huh7
Coronavirus Several BNT162b2 A recent study showed that SARS-CoV-2 RNAs can be reverse-transcribed and integrated into the genome of human cells [25]. Huh7 cells (JCRB Cell Bank, Osaka, Japan) were cultured in 37 °C at 5% CO2 with DMEM medium (HyClone, HYCLSH30243.01) supplemented with 10% (v/v) fetal bovine serum (Sigma-Aldrich, F7524-500ML, Burlington, MA, USA) and 1% (v/v) RNA
Table 1.
Table 1. 1. Introduction
disease 2019 (COVID-19) caused by severe acute respiratory syndrome
coronavirus 2 (SARS-CoV-2) was announced by the World Health
Organization (WHO) as a global pandemic on 11 March 2020, and it emerged
as a devasting health crisis. As of February 2022, COVID-19 has led to
over 430 million reported infection cases and 5.9 million deaths
worldwide [1]. Effective and safe vaccines are urgently needed to reduce the morbidity and mortality rates associated with COVID-19.
vaccines for COVID-19 have been developed, with particular focus on
mRNA vaccines (by Pfizer-BioNTech and Moderna), replication-defective
recombinant adenoviral vector vaccines (by Janssen-Johnson and Johnson,
Astra-Zeneca, Sputnik-V, and CanSino), and inactivated vaccines (by
Sinopharm, Bharat Biotech and Sinovac). The mRNA vaccine has the
advantages of being flexible and efficient in immunogen design and
manufacturing, and currently, numerous vaccine candidates are in various
stages of development and application. Specifically, COVID-19 mRNA
vaccine BNT162b2 developed by Pfizer and BioNTech has been evaluated in
successful clinical trials [2,3,4] and administered in national COVID-19 vaccination campaigns in different regions around the world [5,6,7,8].
is a lipid nanoparticle (LNP)–encapsulated, nucleoside-modified RNA
vaccine (modRNA) and encodes the full-length of SARS-CoV-2 spike (S)
protein, modified by two proline mutations to ensure antigenically
optimal pre-fusion conformation, which mimics the intact virus to elicit
virus-neutralizing antibodies [3].
Consistent with randomized clinical trials, BNT162b2 showed high
efficiency in a wide range of COVID-19-related outcomes in a real-world
setting [5].
Nevertheless, many challenges remain, including monitoring for
long-term safety and efficacy of the vaccine. This warrants further
evaluation and investigations. The safety profile of BNT162b2 is
currently only available from short-term clinical studies. Less common
adverse effects of BNT162b2 have been reported, including pericarditis,
arrhythmia, deep-vein thrombosis, pulmonary embolism, myocardial
infarction, intracranial hemorrhage, and thrombocytopenia [4,9,10,11,12,13,14,15,16,17,18,19,20]. There are also studies that report adverse effects observed in other types of vaccines [21,22,23,24].
To better understand mechanisms underlying vaccine-related adverse
effects, clinical investigations as well as cellular and molecular
analyses are needed.
This gives rise to the question of if this may also occur with
BNT162b2, which encodes partial SARS-CoV-2 RNA. In pharmacokinetics data
provided by Pfizer to European Medicines Agency (EMA), BNT162b2
biodistribution was studied in mice and rats by intra-muscular injection
with radiolabeled LNP and luciferase modRNA. Radioactivity was detected
in most tissues from the first time point (0.25 h), and results showed
that the injection site and the liver were the major sites of
distribution, with maximum concentrations observed at 8–48 h post-dose [26].
Furthermore, in animals that received the BNT162b2 injection,
reversible hepatic effects were observed, including enlarged liver,
vacuolation, increased gamma glutamyl transferase (γGT) levels, and
increased levels of aspartate transaminase (AST) and alkaline
phosphatase (ALP) [26]. Transient hepatic effects induced by LNP delivery systems have been reported previously [27,28,29,30],
nevertheless, it has also been shown that the empty LNP without modRNA
alone does not introduce any significant liver injury [27].
Therefore, in this study, we aim to examine the effect of BNT162b2 on a
human liver cell line in vitro and investigate if BNT162b2 can be
reverse transcribed into DNA through endogenous mechanisms. 2. Materials and Methods
2.1. Cell Culture
Penicillin-Streptomycin (HyClone, SV30010, Logan, UT, USA). For
BNT162b2 treatment, Huh7 cells were seeded with a density of 200,000
cells/well in 24-well plates. BNT162b2 mRNA vaccine (Pfizer BioNTech,
New York, NY, USA) was diluted with sterile 0.9% sodium chloride
injection, USP into a final concentration of 100 μg/mL as described in
the manufacturer’s guideline [31].
BNT162b2 suspension was then added in cell culture media to reach final
concentrations of 0.5, 1.0, or 2.0 μg/mL. Huh7 cells were incubated
with or without BNT162b2 for 6, 24, and 48 h. Cells were washed
thoroughly with PBS and harvested by trypsinization and stored in −80 °C
until further use. 2.2. REAL-TIME RT-QPCR
from the cells was extracted with RNeasy Plus Mini Kit (Qiagen, 74134,
Hilden, Germany) following the manufacturer’s protocol. RT-PCR was
performed using RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, K1622, Waltham, MA, USA) following the manufacturers
protocol. Real-time qPCR was performed using Maxima SYBR Green/ROX qPCR
Master Mix (Thermo Fisher Scientific, K0222, Waltham, MA, USA) with
primers for BNT162b2, LINE-1 and housekeeping genes ACTB and GAPDH (Table 1).
Primer sequences of RT-qPCR and PCR.

Primer sequences of RT-qPCR and PCR.
Target
Sequence
ACTB forward
CCTCGCCTTTGCCGATCC
ACTB reverse
GGATCTTCATGAGGTAGTCAGTC
GAPDH forward
CTCTGCTCCTCCTGTTCGAC
GAPDH reverse
TTAAAAGCAGCCCTGGTGAC
LINE-1 forward
TAACCAATACAGAGAAGTGC
LINE-1 reverse
GATAATATCCTGCAGAGTGT
BNT162b2 forward
CGAGGTGGCCAAGAATCTGA
BNT162b2 reverse
TAGGCTAAGCGTTTTGAGCTG
2.3. Immunofluorescence Staining and Confocal Imaging
Huh7
cells were cultured in eight-chamber slides (LAB-TEK, 154534, Santa
Cruz, CA, USA) with a density of 40,000 cells/well, with or without
BNT162b2 (0.5, 1 or 2 µg/mL) for 6 h. Immunohistochemistry was performed
using primary antibody anti-LINE-1 ORF1p mouse monoclonal antibody
(Merck, 3574308, Kenilworth, NJ, USA), secondary antibody Cy3 Donkey
anti-mouse (Jackson ImmunoResearch, West Grove, PA, USA), and Hoechst
(Life technologies, 34850, Carlsbad, CA, USA), following the protocol
from Thermo Fisher (Waltham, MA, USA). Two images per condition were
taken using a Zeiss LSM 800 and a 63X oil immersion objective, and the
staining intensity was quantified on the individual whole cell area and
the nucleus area on 15 cells per image by ImageJ 1.53c. LINE-1 staining
intensity for the cytosol was calculated by subtracting the intensity of
the nucleus from that of the whole cell. All images of the cells were
assigned a random number to prevent bias. To mark the nuclei (determined
by the Hoechst staining) and the whole cells (determined by the borders
of the LINE-1 fluorescence), the Freehand selection tool was used.
These areas were then measured, and the mean intensity was used to
compare the groups.
2.4. Genomic DNA Purification, PCR Amplification, Agarose Gel Purification, and Sanger Sequencing
Genomic
DNA was extracted from cell pellets with PBND buffer (10 mM Tris-HCl pH
8.3, 50 mM KCl, 2.5 mM MgCl2, 0.45% NP-40, 0.45% Tween-20) according to
protocol described previously [32].
To remove residual RNA from the DNA preparation, RNase (100 µg/mL,
Qiagen, Hilden, Germany) was added to the DNA preparation and incubated
at 37 °C for 3 h, followed by 5 min at 95 °C. PCR was then performed
using primers targeting BNT162b2 (sequences are shown in Table 1),
with the following program: 5 min at 95 °C, 35 cycles of 95 °C for 30
s, 58 °C for 30 s, and 72 °C for 1 min; finally, 72 °C for 5 min and 12
°C for 5 min. PCR products were run on 1.4% (w/v)
agarose gel. Bands corresponding to the amplicons of the expected size
(444 bps) were cut out and DNA was extracted using QIAquick PCR
Purification Kit (Qiagen, 28104, Hilden, Germany), following the
manufacturer’s instructions. The sequence of the DNA amplicon was
verified by Sanger sequencing (Eurofins Genomics, Ebersberg, Germany).
Statistics
Statistical comparisons were performed using two-tailed Student’s t-test and ANOVA. Data are expressed as the mean ± SEM or ± SD. Differences with p < 0.05 are considered significant.
2.5. Ethical Statements
The Huh7 cell line was obtained from Japanese Collection of Research Bioresources (JCRB) Cell Bank.
3. Results
3.1. BNT162b2 Enters Human Liver Cell Line Huh7 Cells at High Efficiency
To
determine if BNT162b2 enters human liver cells, we exposed human liver
cell line Huh7 to BNT162b2. In a previous study on the uptake kinetics
of LNP delivery in Huh7 cells, the maximum biological efficacy of LNP
was observed between 4–7 h [33].
Therefore, in our study, Huh7 cells were cultured with or without
increasing concentrations of BNT162b2 (0.5, 1.0 and 2.0 µg/mL) for 6,
24, and 48 h. RNA was extracted from cells and a real-time quantitative
reverse transcription polymerase chain reaction (RT-qPCR) was performed
using primers targeting the BNT162b2 sequence, as illustrated in Figure 1. The full sequence of BNT162b2 is publicly available [34]
and contains a two-nucleotides cap; 5′- untranslated region (UTR) that
incorporates the 5′ -UTR of a human α-globin gene; the full-length of
SARS-CoV-2 S protein with two proline mutations; 3′-UTR that
incorporates the human mitochondrial 12S rRNA (mtRNR1) segment and human
AES/TLE5 gene segment with two C→U mutations; poly(A) tail. Detailed
analysis of the S protein sequence in BNT162b2 revealed 124 sequences
that are 100% identical to human genomic sequences and three sequences
with only one nucleotide (nt) mismatch in 19–26 nts (Table S1, see Supplementary Materials).
To detect BNT162b2 RNA level, we designed primers with forward primer
located in SARS-CoV-2 S protein regions and reverse primer in 3′-UTR,
which allows detection of PCR amplicon unique to BNT162b2 without
unspecific binding of the primers to human genomic regions.
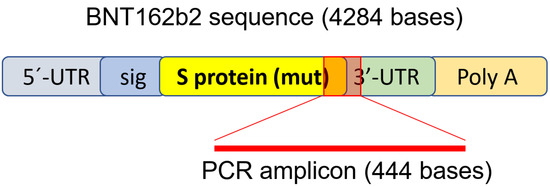
Figure 1.
PCR primer set used to detect mRNA level and reverse-transcription of
BNT162b2. Illustration of BNT162b2 was adapted from previously described
literature [34].
RT-qPCR results showed that Huh7 cells treated
with BNT162b2 had high levels of BNT162b2 mRNA relative to housekeeping
genes at 6, 24, and 48 h (Figure 2, presented in logged 2−ΔΔCT
due to exceptionally high levels). The three BNT162b2 concentrations
led to similar intracellular BNT162b2 mRNA levels at the different time
points, except that the significant difference between 1.0 and 2.0 µg/mL
was observed at 48 h. BNT162b2 mRNA levels were significantly decreased
at 24 h compared to 6 h, but increased again at 48 h.
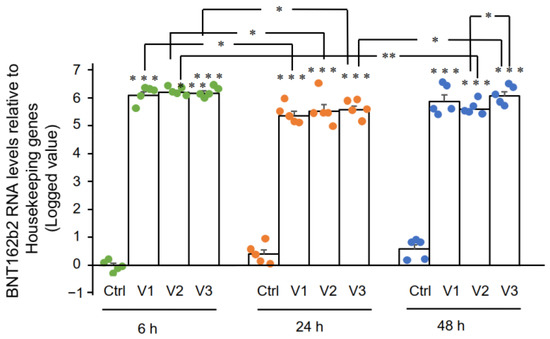
Figure 2.
BNT162b2 mRNA levels in Huh7 cells treated with BNT162b2. Huh7 cells
were treated without (Ctrl) or with 0.5 (V1), 1 (V2), and 2 µg/mL (V3)
of BNT162b2 for 6 (green dots), 24 (orange dots), and 48 h (blue dots).
RNA was purified and qPCR was performed using primers targeting
BNT162b2. RNA levels of BNT162b2 are presented as logged 2−ΔΔCT values relative to house-keeping genes GAPDH and ACTB. Results are from five independent experiments (n = 5). Differences between respective groups were analyzed using two-tailed Student’s t-test. Data are expressed as the mean ± SEM. (* p < 0.05; ** p < 0.01; *** p < 0.001 vs. respective control at each time point, or as indicated).
3.2. Effect of BNT162b2 on Human Endogenous Reverse Transcriptase Long Interspersed Nuclear Element-1 (LINE-1)
Here we examined the effect of BNT162b2 on LINE-1
gene expression. RT-qPCR was performed on RNA purified from Huh7 cells
treated with BNT162b2 (0, 0.5, 1.0, and 2.0 µg/mL) for 6, 24, and 48 h,
using primers targeting LINE-1. Significantly increased LINE-1 expression compared to control was observed at 6 h by 2.0 µg/mL BNT162b2, while lower BNT162b2 concentrations decreased LINE-1 expression at all time points (Figure 3).
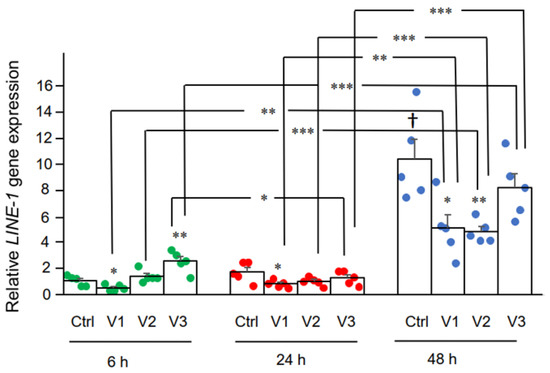
Figure 3.
LINE-1 mRNA levels in Huh7 cells
treated with BNT162b2. Huh7 cells were treated without (Ctrl) or with
0.5 (V1), 1 (V2), and 2 µg/mL (V3) of BNT162b2 for 6 (green dots), 24
(red dots), and 48 h (blue dots). RNA was purified and qPCR was
performed using primers targeting LINE-1. RNA levels of LINE-1 are presented as 2−ΔΔCT values relative to house-keeping genes GAPDH and ACTB. Results are from five independent experiments (n = 5). Differences between respective groups were analyzed using two-tailed Student’s t-test. Data are expressed as the mean ± SEM. (* p < 0.05; ** p < 0.01; *** p < 0.001 vs. respective control at each time point, or as indicated; † p < 0.05 vs. 6 h-Ctrl).
Next, we studied the effect of BNT162b2 on
LINE-1 protein level. The full-length LINE-1 consists of a 5′
untranslated region (UTR), two open reading frames (ORFs), ORF1 and
ORF2, and a 3′UTR, of which ORF1 is an RNA binding protein with
chaperone activity. The retrotransposition activity of LINE-1 has been
demonstrated to involve ORF1 translocation to the nucleus [35].
Huh7 cells treated with or without BNT162b2 (0.5, 1.0 and 2.0 µg/mL)
for 6 h were fixed and stained with antibodies binding to LINE-1 ORF1p,
and DNA-specific probe Hoechst for visualization of cell nucleus (Figure 4a).
Quantification of immunofluorescence staining intensity showed that
BNT162b2 increased LINE-1 ORF1p protein levels in both the whole cell
area and nucleus at all concentrations tested (Figure 4b–d).
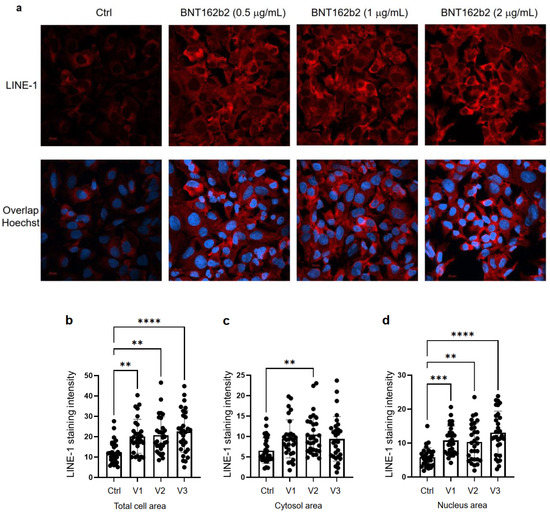
Figure 4.
Immunohistochemistry of Huh7 cells treated with BNT162b2 on LINE-1
protein distribution. Huh7 cells were treated without (Ctrl) or with
0.5, 1, and 2 µg/mL of BNT162b2 for 6 h. Cells were fixed and stained
with antibodies binding to LINE-1 ORF1p (red) and DNA-specific probe
Hoechst for visualization of cell nucleus (blue). (a) Representative images of LINE-1 expression in Huh7 cells treated with or without BNT162b2. (b–d) Quantification of LINE-1 protein in whole cell area (b), cytosol (c), and nucleus (d).
All data were analyzed using One-Way ANOVA, and graphs were created
using GraphPad Prism V 9.2. All data is presented as mean ± SD (** p < 0.01; *** p < 0.001; **** p < 0.0001 as indicated).
3.3. Detection of Reverse Transcribed BNT162b2 DNA in Huh7 Cells
A previous study has shown that entry of LINE-1 protein into the nucleus is associated with retrotransposition [35].
In the immunofluorescence staining experiment described above,
increased levels of LINE-1 in the nucleus were observed already at the
lowest concentration of BNT162b2 (0.5 µg/mL). To examine if BNT162b2 is
reversely transcribed into DNA when LINE-1 is elevated, we purified
genomic DNA from Huh7 cells treated with 0.5 µg/mL of BNT162b2 for 6,
24, and 48 h. Purified DNA was treated with RNase to remove RNA and
subjected to PCR using primers targeting BNT162b2, as illustrated in Figure 1. Amplified DNA fragments were then visualized by electrophoresis and gel-purified (Figure 5).
BNT162b2 DNA amplicons were detected in all three time points (6, 24,
and 48 h). Sanger sequencing confirmed that the DNA amplicons were
identical to the BNT162b2 sequence flanked by the primers (Table 2).
To ensure that the DNA amplicons were derived from DNA but not BNT162b2
RNA, we also performed PCR on RNA purified from Huh7 cells treated with
0.5 µg/mL BNT162b2 for 6 h, with or without RNase treatment (Ctrl 5 and
6 in Figure 5), and no amplicon was detected in the RNA samples subjected to PCR.
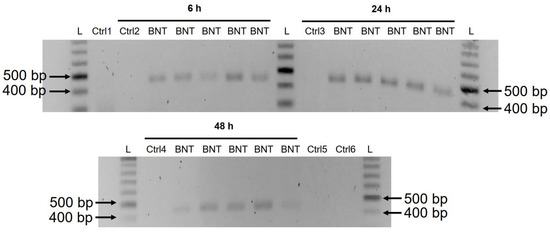
Figure 5.
Detection of DNA amplicons of BNT162b2 in Huh7 cells treated with
BNT162b2. Huh7 cells were treated without (Ctrl) or with 0.5 µg/mL of
BNT162b2 for 6, 24, and 48 h. Genomic DNA was purified and digested with
100 µg/mL RNase. PCR was run on all samples with primers targeting
BNT162b2, as shown in Figure 1 and Table 1.
DNA amplicons (444 bps) were visualized on agarose gel. BNT: BNT162b2;
L: DNA ladder; Ctrl1: cultured Huh7 cells; Ctrl2: Huh7 cells without
BNT162b2 treatment collected at 6 h; Ctrl3: Huh7 cells without BNT162b2
treatment collected at 24 h; Ctrl4: Huh7 cells without BNT162b2
treatment collected at 48 h; Ctrl5: RNA from Huh7 cells treated with 0.5
µg/mL of BNT162b2 for 6 h; Ctrl6: RNA from Huh7 cells treated with 0.5
µg/mL of BNT162b2 for 6 h, digested with RNase.
Table 2.
Sanger sequencing result of the BNT162b2 amplicon.
Table 2.
Sanger sequencing result of the BNT162b2 amplicon.
| CGAGGTGGCCAAGAATCTGAACGAGAGCCTGATCGACCTGCAAGAACTGGGGAAGT ACGAGCAGTACATCAAGTGGCCCTGGTACATCTGGCTGGGCTTTATCGCCGGACTGATTG CCATCGTGATGGTCACAATCATGCTGTGTTGCATGACCAGCTGCTGTAGCTGCCTGAAGG GCTGTTGTAGCTGTGGCAGCTGCTGCAAGTTCGACGAGGACGATTCTGAGCCCGTGCTGA |
| AGGGCGTGAAACTGCACTACACATGATGACTCGAGCTGGTACTGCATGCACGCAATGCTA GCTGCCCCTTTCCCGTCCTGGGTACCCCGAGTCTCCCCCGACCTCGGGTCCCAGGTATGC TCCCACCTCCACCTGCCCCACTCACCACCTCTGCTAGTTCCAGACACCTCCCAAGCACGC AGCAATGCAGCTCAAAACGCTTAGCCTA |
4. Discussion
In
this study we present evidence that COVID-19 mRNA vaccine BNT162b2 is
able to enter the human liver cell line Huh7 in vitro. BNT162b2 mRNA is
reverse transcribed intracellularly into DNA as fast as 6 h after
BNT162b2 exposure. A possible mechanism for reverse transcription is
through endogenous reverse transcriptase LINE-1, and the nucleus protein
distribution of LINE-1 is elevated by BNT162b2.
Intracellular accumulation of LNP in hepatocytes has been demonstrated in vivo [36].
A preclinical study on BNT162b2 showed that BNT162b2 enters the human
cell line HEK293T cells and leads to robust expression of BNT162b2
antigen [37].
Therefore, in this study, we first investigated the entry of BNT162b2
in the human liver cell line Huh7 cells. The choice of BNT162b2
concentrations used in this study warrants explanation. BNT162b2 is
administered as a series of two doses three weeks apart, and each dose
contains 30 µg of BNT162b2 in a volume of 0.3 mL, which makes the local
concentration at the injection site at the highest 100 µg/mL [31].
A previous study on mRNA vaccines against H10N8 and H7N9 influenza
viruses using a similar LNP delivery system showed that the mRNA vaccine
can distribute rather nonspecifically to several organs such as liver,
spleen, heart, kidney, lung, and brain, and the concentration in the
liver is roughly 100 times lower than that of the intra-muscular
injection site [38].
In the assessment report on BNT162b2 provided to EMA by Pfizer, the
pharmacokinetic distribution studies in rats demonstrated that a
relatively large proportion (up to 18%) of the total dose distributes to
the liver [26].
We therefore chose to use 0.5, 1, and 2 μg/mL of vaccine in our
experiments on the liver cells. However, the effect of a broader range
of lower and higher concentrations of BNT162b2 should also be verified
in future studies.
In the current study, we
employed a human liver cell line for in vitro investigation. It is worth
investigating if the liver cells also present the vaccine-derived
SARS-CoV-2 spike protein, which could potentially make the liver cells
targets for previously primed spike protein reactive cytotoxic T cells.
There has been case reports on individuals who developed autoimmune
hepatitis [39]
after BNT162b2 vaccination. To obtain better understanding of the
potential effects of BNT162b2 on liver function, in vivo models are
desired for future studies.
In the BNT162b2 toxicity report, no genotoxicity nor carcinogenicity studies have been provided [26].
Our study shows that BNT162b2 can be reverse transcribed to DNA in
liver cell line Huh7, and this may give rise to the concern if
BNT162b2-derived DNA may be integrated into the host genome and affect
the integrity of genomic DNA, which may potentially mediate genotoxic
side effects. At this stage, we do not know if DNA reverse transcribed
from BNT162b2 is integrated into the cell genome. Further studies are
needed to demonstrate the effect of BNT162b2 on genomic integrity,
including whole genome sequencing of cells exposed to BNT162b2, as well
as tissues from human subjects who received BNT162b2 vaccination.
Human
autonomous retrotransposon LINE-1 is a cellular endogenous reverse
transcriptase and the only remaining active transposon in humans, able
to retrotranspose itself and other nonautonomous elements [40,41], and ~17% of the human genome are comprised of LINE-1 sequences [42]. The nonautonomous Alu
elements, short, interspersed nucleotide elements (SINEs),
variable-number-of-tandem-repeats (VNTR), as well as cellular
mRNA-processed pseudogenes, are retrotransposed by the LINE-1
retrotransposition proteins working in trans [43,44].
A recent study showed that endogenous LINE-1 mediates reverse
transcription and integration of SARS-CoV-2 sequences in the genomes of
infected human cells [25]. Furthermore, expression of endogenous LINE-1 is often increased upon viral infection, including SARS-CoV-2 infection [45,46,47]. Previous studies showed that LINE-1 retrotransposition activity is regulated by RNA metabolism [48,49], DNA damage response [50], and autophagy [51]. Efficient retrotransposition of LINE-1 is often associated with cell cycle and nuclear envelope breakdown during mitosis [52,53], as well as exogenous retroviruses [54,55],
which promotes entrance of LINE-1 into the nucleus. In our study, we
observed increased LINE-1 ORF1p distribution as determined by
immunohistochemistry in the nucleus by BNT162b2 at all concentrations
tested (0.5, 1, and 2 μg/mL), while elevated LINE-1
gene expression was detected at the highest BNT162b2 concentration (2
μg/mL). It is worth noting that gene transcription is regulated by
chromatin modifications, transcription factor regulation, and the rate
of RNA degradation, while translational regulation of protein involves
ribosome recruitment on the initiation codon, modulation of peptide
elongation, termination of protein synthesis, or ribosome biogenesis.
These two processes are controlled by different mechanisms, and
therefore they may not always show the same change patterns in response
to external challenges. The exact regulation of LINE-1 activity in
response to BNT162b2 merits further study.
The
cell model that we used in this study is a carcinoma cell line, with
active DNA replication which differs from non-dividing somatic cells. It
has also been shown that Huh7 cells display significant different gene
and protein expression including upregulated proteins involved in RNA
metabolism [56].
However, cell proliferation is also active in several human tissues
such as the bone marrow or basal layers of epithelia as well as during
embryogenesis, and it is therefore necessary to examine the effect of
BNT162b2 on genomic integrity under such conditions. Furthermore,
effective retrotransposition of LINE-1 has also been reported in
non-dividing and terminally differentiated cells, such as human neurons [57,58].
The
Pfizer EMA assessment report also showed that BNT162b2 distributes in
the spleen (<1.1%), adrenal glands (<0.1%), as well as low and
measurable radioactivity in the ovaries and testes (<0.1%) [26].
Furthermore, no data on placental transfer of BNT162b2 is available
from Pfizer EMA assessment report. Our results showed that BNT162b2 mRNA
readily enters Huh7 cells at a concentration (0.5 µg/mL) corresponding
to 0.5% of the local injection site concentration, induce changes in
LINE-1 gene and protein expression, and within 6 h, reverse
transcription of BNT162b2 can be detected. It is therefore important to
investigate further the effect of BNT162b2 on other cell types and
tissues both in vitro and in vivo.
5. Conclusions
Our
study is the first in vitro study on the effect of COVID-19 mRNA
vaccine BNT162b2 on human liver cell line. We present evidence on fast
entry of BNT162b2 into the cells and subsequent intracellular reverse
transcription of BNT162b2 mRNA into DNA.
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/cimb44030073/s1. M.A., This Not applicable. Not applicable. All data supporting the findings of this study are available within the article and supporting information. The The authors declare no conflict of interest. Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. Supplementary Materials
Author Contributions
F.O.F., D.Y., M.B. and C.L. performed in vitro experiments. M.A. and
F.O.F. performed data analysis. M.R. and Y.D.M. contributed to the
implementation of the research, designed, and supervised the study.
Y.D.M. wrote the paper with input from all authors. All authors have
read and agreed to the published version of the manuscript.Funding
study was supported by the Swedish Research Council (SRC), Strategic Research
Area Exodiab, Dnr 2009-1039, the Swedish Government Fund for Clinical
Research (ALF) and the foundation of Skåne University Hospital.Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
authors thank Sven Haidl, Maria Josephson, Enming Zhang, Jia-Yi Li,
Caroline Haikal, and Pradeep Bompada for their support to this study.Conflicts of Interest
References
M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.;
Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II
study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef] [PubMed]
E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.;
Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et
al. Safety and Immunogenicity of Two RNA-Based COVID-19 Vaccine
Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart,
S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al.
Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
R.J.; Hall, J.A.; Zaidi, A.; Andrews, N.J.; Dunbar, J.K.; Dabrera, G.
Effect of Vaccination on Household Transmission of SARS-CoV-2 in
England. N. Engl. J. Med. 2021, 385, 759–760. [Google Scholar] [CrossRef]
A.A.; Omer, S.B.; Yan, P.; Shaikh, O.S.; Mayr, F.B. SARS-CoV-2 Vaccine
Effectiveness in a High-Risk National Population in a Real-World
Setting. Ann. Intern. Med. 2021, 174, 1404–1408. [Google Scholar] [CrossRef]
N.; Barda, N.; Kepten, E.; Miron, O.; Perchik, S.; Katz, M.A.; Hernan,
M.A.; Lipsitch, M.; Reis, B.; Balicer, R.D. BNT162b2 mRNA Covid-19
Vaccine in a Nationwide Mass Vaccination Setting. N. Engl. J. Med. 2021, 384, 1412–1423. [Google Scholar] [CrossRef]
H.; Shilo, S.; Meir, T.; Gorfine, M.; Shalit, U.; Segal, E. COVID-19
dynamics after a national immunization program in Israel. Nat. Med. 2021, 27, 1055–1061. [Google Scholar] [CrossRef]
B.E.; Shen, J.Y.; Lim, X.R.; Tu, T.M.; Chang, C.C.R.; Khin, H.S.W.;
Koh, J.S.; Rao, J.P.; Lau, S.L.; Tan, G.B.; et al. Cerebral venous
thrombosis post BNT162b2 mRNA SARS-CoV-2 vaccination: A black swan
event. Am. J. Hematol. 2021, 96, E357–E361. [Google Scholar] [CrossRef]
K.F.; Ammirati, E.; Adler, E.D.; Cooper, L.T., Jr.; Hong, K.N.;
Saponara, G.; Couri, D.; Cereda, A.; Procopio, A.; Cavalotti, C.; et al.
Myocarditis After BNT162b2 and mRNA-1273 Vaccination. Circulation 2021, 144, 506–508. [Google Scholar] [CrossRef]
C.; Klaser, K.; May, A.; Polidori, L.; Capdevila, J.; Louca, P.; Sudre,
C.H.; Nguyen, L.H.; Drew, D.A.; Merino, J.; et al. Vaccine side-effects
and SARS-CoV-2 infection after vaccination in users of the COVID
Symptom Study app in the UK: A prospective observational study. Lancet Infect. Dis. 2021, 21, 939–949. [Google Scholar] [CrossRef]
T.; Titze, U.; Kulamadayil-Heidenreich, N.S.A.; Glombitza, S.; Tebbe,
J.J.; Rocken, C.; Schulz, B.; Weise, M.; Wilkens, L. First case of
postmortem study in a patient vaccinated against SARS-CoV-2. Int. J. Infect. Dis. 2021, 107, 172–175. [Google Scholar] [CrossRef] [PubMed]
R.A.K.; Janagama, R.; Peruru, S.; Malayala, S.V. Side effects of
BNT162b2 mRNA COVID-19 vaccine: A randomized, cross-sectional study with
detailed self-reported symptoms from healthcare workers. Int. J. Infect. Dis. 2021, 106, 376–381. [Google Scholar] [CrossRef] [PubMed]
C.; Piccolo, V.; Pace, G.; Romano, I.; Argenziano, G.; Bassi, A.
Purpuric lesions on the eyelids developed after BNT162b2 mRNA COVID-19
vaccine: Another piece of SARS-CoV-2 skin puzzle? J. Eur. Acad. Dermatol. Venereol. 2021, 35, e543–e545. [Google Scholar] [CrossRef]
E.J.; Cines, D.B.; Gernsheimer, T.; Kessler, C.; Michel, M.; Tarantino,
M.D.; Semple, J.W.; Arnold, D.M.; Godeau, B.; Lambert, M.P.; et al.
Thrombocytopenia following Pfizer and Moderna SARS-CoV-2 vaccination. Am. J. Hematol. 2021, 96, 534–537. [Google Scholar] [CrossRef]
Y.; Kenig, A.; Tsemach-Toren, T.; Amer, R.; Rubin, L.; Hershkovitz, Y.;
Kharouf, F. Autoimmune phenomena following SARS-CoV-2 vaccination. Int. Immunopharmacol. 2021, 99, 107970. [Google Scholar] [CrossRef]
B.B.; Kohli, U.; Ramachandran, P.; Nguyen, H.H.; Greil, G.; Hussain,
T.; Tandon, A.; Kane, C.; Avula, S.; Duru, C.; et al. Myopericarditis
following mRNA COVID-19 Vaccination in Adolescents 12 through 18 Years
of Age. J. Pediatr. 2021, 238, 26–32.e1. [Google Scholar] [CrossRef]
S.; Garner, C.D.; Krutsch, K.; Hale, T.W. Maternal and Child Symptoms
Following COVID-19 Vaccination Among Breastfeeding Mothers. Breastfeed. Med. 2021, 16, 702–709. [Google Scholar] [CrossRef]
N.; Dagan, N.; Ben-Shlomo, Y.; Kepten, E.; Waxman, J.; Ohana, R.;
Hernan, M.A.; Lipsitch, M.; Kohane, I.; Netzer, D.; et al. Safety of the
BNT162b2 mRNA Covid-19 Vaccine in a Nationwide Setting. N. Engl. J. Med. 2021, 385, 1078–1090. [Google Scholar] [CrossRef]
L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.;
Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy
and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn,
B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al.
Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19.
N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
B.; Ciobica, A.; Ilie, O.D.; Maftei, R.; Ilea, C. Mini-Review
Discussing the Reliability and Efficiency of COVID-19 Vaccines. Diagnostics 2021, 11, 579. [Google Scholar] [CrossRef]
L.; Richards, A.; Barrasa, M.I.; Hughes, S.H.; Young, R.A.; Jaenisch,
R. Reverse-transcribed SARS-CoV-2 RNA can integrate into the genome of
cultured human cells and can be expressed in patient-derived tissues. Proc. Natl. Acad. Sci. USA 2021, 118, e2105968118. [Google Scholar] [CrossRef] [PubMed]
H.; Takata, N.; Sakurai, Y.; Yoshida, T.; Inoue, T.; Tamagawa, S.;
Nakai, Y.; Tange, K.; Yoshioka, H.; Maeki, M.; et al. Delivery of
Oligonucleotides Using a Self-Degradable Lipid-Like Material. Pharmaceutics 2021, 13, 544. [Google Scholar] [CrossRef]
M.; Senn, J.J.; Lynn, A.; Laska, M.; Smith, M.; Platz, S.J.; Bolen, J.;
Hoge, S.; Bulychev, A.; Jacquinet, E.; et al. Safety Evaluation of
Lipid Nanoparticle-Formulated Modified mRNA in the Sprague-Dawley Rat
and Cynomolgus Monkey. Vet. Pathol. 2018, 55, 341–354. [Google Scholar] [CrossRef]
Y.; Matsui, H.; Yamamoto, N.; Sato, R.; Munakata, T.; Kohara, M.;
Harashima, H. Highly specific delivery of siRNA to hepatocytes
circumvents endothelial cell-mediated lipid nanoparticle-associated
toxicity leading to the safe and efficacious decrease in the hepatitis B
virus. J. Control. Release 2017, 266, 216–225. [Google Scholar] [CrossRef]
J.D.; Yu, Z.; Liu, J.Y.; Rele, S.M.; Liang, Y.; Zeidan, R.K.;
Kornbrust, D.J.; Davis, M.E. Administration in non-human primates of
escalating intravenous doses of targeted nanoparticles containing
ribonucleotide reductase subunit M2 siRNA. Proc. Natl. Acad. Sci. USA 2007, 104, 5715–5721. [Google Scholar] [CrossRef]
A.; Munson, M.J.; Liu, K.; Idstrom, A.; Barriga, H.M.; Tabaei, S.;
Aliakbarinodehi, N.; Ojansivu, M.; Lubart, Q.; Doutch, J.J.; et al. Time
evolution of PEG-shedding and serum protein coronation determines the
cell uptake kinetics and delivery of lipid nanoparticle. bioRxiv 2021. [Google Scholar] [CrossRef]
P.; Wudzinska, A.; Sun, X.; Andrade, J.; Nayak, S.; Kahler, D.J.;
Badri, S.; LaCava, J.; Ueberheide, B.; Yun, C.Y.; et al. LINE-1 protein
localization and functional dynamics during the cell cycle. Elife 2018, 7, e30058. [Google Scholar] [CrossRef]
Y.; Kinami, Y.; Hashiba, K.; Harashima, H. Different kinetics for the
hepatic uptake of lipid nanoparticles between the apolipoprotein E/low
density lipoprotein receptor and the
N-acetyl-d-galactosamine/asialoglycoprotein receptor pathway. J. Control. Release 2020, 322, 217–226. [Google Scholar] [CrossRef]
A.B.; Kanevsky, I.; Che, Y.; Swanson, K.A.; Muik, A.; Vormehr, M.;
Kranz, L.M.; Walzer, K.C.; Hein, S.; Guler, A.; et al. BNT162b vaccines
protect rhesus macaques from SARS-CoV-2. Nature 2021, 592, 283–289. [Google Scholar] [CrossRef] [PubMed]
K.; Senn, J.J.; Yuzhakov, O.; Bulychev, A.; Brito, L.A.; Hassett, K.J.;
Laska, M.E.; Smith, M.; Almarsson, O.; Thompson, J.; et al. Preclinical
and Clinical Demonstration of Immunogenicity by mRNA Vaccines against
H10N8 and H7N9 Influenza Viruses. Mol. Ther. 2017, 25, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
F.; Al Diffalha, S.; Dean, M.; Fettig, D.M. Autoimmune hepatitis
developing after coronavirus disease 2019 (COVID-19) vaccine: Causality
or casualty? J. Hepatol. 2021, 75, 222–224. [Google Scholar] [CrossRef]
E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.;
Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing
and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
E.M.; Goodier, J.L.; Zhang, Y.; Kazazian, H.H., Jr. SVA elements are
nonautonomous retrotransposons that cause disease in humans. Am. J. Hum. Genet. 2003, 73, 1444–1451. [Google Scholar] [CrossRef]
R.B.; Song, H.; Xu, Y.; Garrison, K.E.; Buzdin, A.A.; Anwar, N.;
Hunter, D.V.; Mujib, S.; Mihajlovic, V.; Martin, E.; et al. LINE-1
retrotransposable element DNA accumulates in HIV-1-infected cells. J. Virol. 2013, 87, 13307–13320. [Google Scholar] [CrossRef]
M.G.; Langlois, R.A.; Shen, S.S. Virus-induced transposable element
expression up-regulation in human and mouse host cells. Life Sci. Alliance 2020, 3, e201900536. [Google Scholar] [CrossRef] [PubMed]
L.; Taylor, M.S.; O’Donnell, K.A.; Boeke, J.D. Poly(A) binding protein
C1 is essential for efficient L1 retrotransposition and affects L1 RNP
formation. Mol. Cell Biol. 2012, 32, 4323–4336. [Google Scholar] [CrossRef]
G.; Streva, V.A.; Derbes, R.S.; Wijetunge, M.I.; Neeland, M.; White,
T.B.; Belancio, V.P.; Roy-Engel, A.M.; Deininger, P.L. The Nucleotide
Excision Repair Pathway Limits L1 Retrotransposition. Genetics 2017, 205, 139–153. [Google Scholar] [CrossRef]
H.; Chitiprolu, M.; Gagnon, D.; Meng, L.; Perez-Iratxeta, C.; Lagace,
D.; Gibbings, D. Autophagy supports genomic stability by degrading
retrotransposon RNA. Nat. Commun. 2014, 5, 5276. [Google Scholar] [CrossRef]
Y.; Mates, L.; Ivics, Z.; Izsvak, Z.; Martin, S.L.; An, W. Cell
division promotes efficient retrotransposition in a stable L1 reporter
cell line. Mob. DNA 2013, 4, 10. [Google Scholar] [CrossRef]
J.; Wang, X.; Lyu, L.; Jiang, H.; Zhu, H.J. Comparison of protein
expression between human livers and the hepatic cell lines HepG2, Hep3B,
and Huh7 using SWATH and MRM-HR proteomics: Focusing on
drug-metabolizing enzymes. Drug Metab. Pharmacokinet. 2018, 33, 133–140. [Google Scholar] [CrossRef] [PubMed]
S.; Seleme, M.C.; Soifer, H.S.; Perez, J.L.; Moran, J.V.; Kazazian,
H.H., Jr.; Kasahara, N. L1 retrotransposition in nondividing and primary
human somatic cells. Proc. Natl. Acad. Sci. USA 2006, 103, 8036–8041. [Google Scholar] [CrossRef] [PubMed]
A.; Widmann, T.J.; Heras, S.R.; Ayllon, V.; Sanchez, L.;
Benkaddour-Boumzaouad, M.; Munoz-Lopez, M.; Rubio, A.; Amador-Cubero,
S.; Blanco-Jimenez, E.; et al. Engineered LINE-1 retrotransposition in
nondividing human neurons. Genome Res. 2017, 27, 335–348. [Google Scholar] [CrossRef] [PubMed]
©
2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is
an open access article distributed under the terms and conditions of
the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
==========================================
Meer over het farmaceutische maffia lid Pfizer: ‘Pfizer CEO: Israëliërs zijn proefkonijn voor testen van vaccin‘ (!!!!) De CEO van Pfizer die zonder blikken of blozen aangeeft waar het om gaat als men gevaccineerd is……. Israël
waar intussen velen al spijt hebben zich te hebben laten
vaccineren….. Intussen heeft Pfizer afgezien van een contract met
India, daar het land extra garanties wilde over de veiligheid en daar
zag Pfizer geen brood in…… India een land met een bevolking van 1,3
miljard mensen, ofwel daar was genoeg winst te behalen voor Pfizer; moet
je nagaan….. Er is nog een groot land dat extra veiligheid eiste,
waarop Pfizer ook de stekker uit die onderhandelingen trok….. Als je
hier naar zoekt op het net, maakt niet uit op wat voor manier, vind je
hier niets over terug, tekenend……..
‘Pfizer CEO calls people who spread vaccine misinformation ‘criminals’‘
Een lid van de farmaceutische maffia die anderen aanwijst als
misdadigers….. ha! ha! ha!
‘COVID-19: FDA vraagt
om volledig rapport over toelating van het Pfizer vaccin tot 2076 geheim
te houden >> klokkenluider ontslagen‘ (!!!!) Ook die zaak
wekt ‘veel vertrouwen’ bij de bevolking, vertrouwen in het vaccin, de
Biden administratie en overheidsinstanties als de FDA……. Zoveel
vertrouwen als het volgende:
‘COVID-19-vaccins: Pfizer maakt misbruik van wanhopige regeringen‘
‘COVID-19: Pfizer adviseert derde vaccinatie, omzet van dit bedrijf op deze vaccins in 2021 al meer dan 20 miljard dollar…….‘ (een bericht van 9 juli jl. 2021…..)
‘Pfizer Coronavirus vaccin: wat je niet wordt verteld over de vaccins tegen COVID-19‘
In augustus 2020 deed Pfizer CEO Bourla 62% van z’n aandelen Pfizer van
de hand, blijkbaar heeft hij er geen vertrouwen in dat de beloften van
overheden dat het bedrijf niet vervolgd zal worden voor ernstige
bijwerkingen ook zullen worden geëerbiedigd door rechters mocht men het
bedrijf aanklagen voor doden als gevolg van vaccinatie dan wel voor ernstige (chronische)
bijwerkingen……
——————————————
Zie voorts: ‘Aantal COVID-doden in VS en GB zwaar gemanipuleerd‘
‘VVD trapt werklozen en bijstandsgerechtigden keihard in het kruis‘
‘COVID-maatregelen: een open brief aan Ernst Kuipers‘
‘Aartsbisschop Carlo Maria Figanò: “Twee jaar lang zijn we nu getuige van een globale staatsgreep”‘
(door de Coronamaatregelen) Overigens onbegrijpelijk dat deze
aartsbisschop zich in de bijgevoegde video alleen tot katholieken
richt……
‘COVID-19: onderzoeken bevestigen dat natuurlijke verkregen immuniteit meer dan voldoende is‘ (!!!!) Op 15 februari 2022, werd bekendgemaakt dat natuurlijke
immuniteit veel langer standhoudt dan die middels een vaccin verkregen
en dat niet misselijk: volgens onderzoekers in Italië zelfs 18 maanden!! >> ‘Natural Immunity Lasts for at Least 18 Months: Study‘ En wat
heeft Nederland gedaan?Juist, die periode van natuurlijke immuniteit
teruggebracht naar een half jaar!! Gelukkig zijn we voorlopig af van die
belachelijke beslissing!!
‘Onverdeeld Open: teken het manifest om het Coronatoegangsbewijs onmiddellijk af te schaffen‘
‘Hans Teeuwen: “Geen Coronapas”‘
‘Intrekken uitzendlicentie door Rusland voor Deutsche Welle ‘is een klap in het gezicht van het Duitse volk’‘
RT Deutsch dat werd platgelegd in Duitsland wordt o.a. verweten
complottheorieën over COVID te brengen, dit door het uitzenden van een
gesprek met een paar COVID-vaccinatieweigeraars……. (te zot voor
woorden!!) Oh, en Deutsche Welle is een Duitse staatsomroep die westerse
propaganda uitzond in Rusland, iets
waarover men niet lult in het westen en als men er al over spreeekt
wordt er onomwonden gezegd dat dit soort westerse platforms
onafhankelijk zijn….. ha! ha! ha! ha! ha! ha! ha!
‘EU
medicijnregelgever EMA en de WHO roepen op te stoppen met COVID
boosters vanwege aantasting immuunsysteem en falen bij bestrijding
Coronavirus‘ (!!!!) zie wat dit betreft ook:
‘COVID-19 vaccin zou het immuunsysteem aantasten…….‘ (!!!!)
Benieuwd ook of de vandaag (het is terwijl ik dit schrijf 11 januari 2022 /////) overleden
David Sassoli, voorzitter van het EU parlement, 3 dubbel gevaccineerd
was, daar ook hij een afwijking had aan het immuunsysteem……
‘COVID-19: IC verpleegkundige OLVG slaakt noodkreet‘
‘COVID-19: het bewijs voor Fauci’s oorlog tegen de wetenschap‘ Zie wat dat betreft ook: ‘Great Barrington Declaration tegen overmatige en zelfs gevaarlijke Coronamaatregelen‘
‘COVID-19 ‘booster’:
Israël gaat over op een tweede ‘boostervaccinatie’ vanwege de
omikronvariant, terwijl men in het westen stelt dat de eerste booster
voldoende beschermt tegen die variant‘
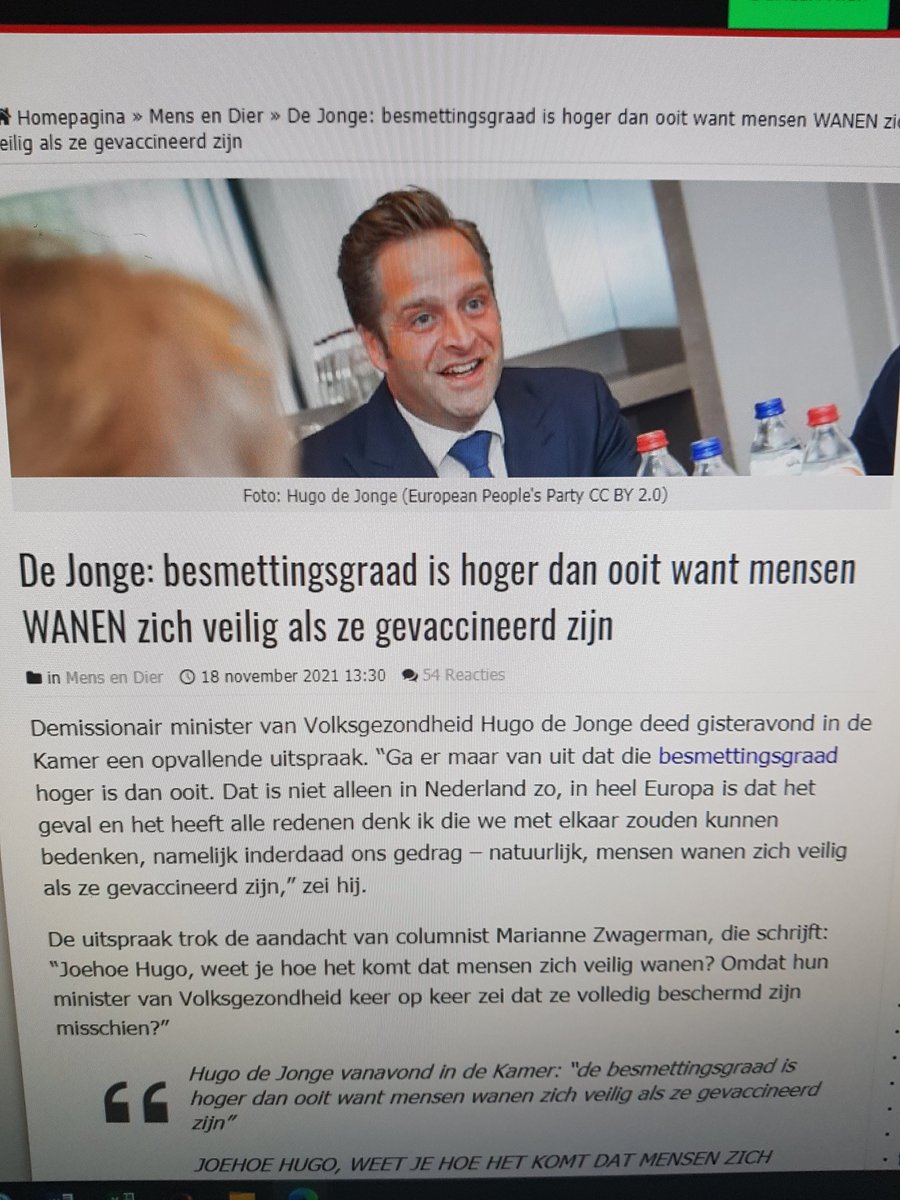
En dit nadat de ex-minister van Volksgezondheid, CDA blunderkont Hugo
de Jonge vorig jaar liet weten dat de eerste 2 vaccinaties voldoende
bescherming zouden bieden tegen nieuwe varianten, om daarna op te merken
dat mensen die gevaccineerd zijn niet moeten denken dat ze veilig
zijn….. ha! ha! ha! ha! (zie de afbeelding met tekst hierboven, in het blok met links naar Pfizer berichten )
Hier nog wat meer links naar ‘oudere berichten’: ‘COVID-19:
Gezondheidsraad houdt zich niet aan de regels van de Raad van Europa:
discriminatie van niet-gevaccineerden is verboden‘ (!!!!)
‘Besmette gevaccineerden houden ‘gewoon’ toegang tot openbare gelegenheden‘
‘Coronavirus:
Diederik Gommers (IC arts) zou het mooi vinden dat mensen worden
ingeënt met het AZ vaccin nadat ze tekenden voor het risico dat ze
lopen…..‘ Dan te bedenken dat zelfs de farmaceuten hun eigen vaccin niet vertrouwen, zie wat dat betreft:
‘Coronavaccinatie ook als je weet dat de fabrikant ernstige bijwerkingen niet uitsluit?‘ En zie:
‘Coronavaccinatie
in vergelijking met het kopen van een huis (waar farmaceuten elke
verantwoordelijkheid voor bijwerkingen afwijzen!!)‘ (en zie de links in dat bericht)
‘Coronavirus: paniek over de Deltavariant is onterecht en zwaar overdreven‘ (!!!!)
En zie de links in dat bericht naar artikelen over de PCR test en de
bezuinigingen op de gezondheidszorg >> die bezuinigingen zijn de
reden voor de
Coronamaatregelen, die werden immers genomen daar er te weinig
ziekenhuisbedden, IC bedden en verpleegkundigen waren. Zoals ook in het bericht hierboven al gemeld: het aantal
verpleegkundigen is als de bedden sterk teruggelopen door wanbeleid van
de afbraakkabinetten Rutte 1, 2 en 3….. Zo waren er bij aantreden van
Rutte 1 nog 2.200 IC bedden, dat was bij aanvang van de Coronacrisis
teruggebracht naar 1.100 bedden….. Dus als je sterk bent
benadeeld door de Coronamaatregelen, weet je wie hiervoor
verantwoordelijk zijn!! Zie wat betreft dat bericht ook: ‘Coronavirus: artsen willen via rechtszaak tegen de staat COVID-19 van A-lijst krijgen, daar het een griepvirus zou zijn ‘De video die oorspronkelijk in het artikel was te zien is
gecensureerd, tja je wilt natuurlijk niet dat de bevolking begrijpt
hoezeer men werd en wordt belazerd……
‘Met COVID-19 besmette zorgmedewerkers mogen ondanks symptomen gewoon doorwerken…….‘
‘Coronavirus: Fort Detrick eindelijk als verdachte van oorsprong aangemerkt‘
‘Vaccinatie tegen Spaanse Griep: miljoenen doden‘
‘COVID vaccinmakers: hoe betrouwbaar zijn deze farmaceuten‘ (en zie de links in dat bericht)
‘COVID-19: gezonde mensen maken elkaar niet ziek‘
‘Coronabeslommeringen: ook na vaccinatie kan je het virus doorgeven, plus de zoveelste blunder van CDA minister de Jonge‘
Het na dit bericht gebrachte nieuws dat je niet besmettelijk bent na
vaccinatie, is achteraf gebeleken grote onzin te zijn en zou gebaseerd
zijn op een
foute interpretatie van een bericht uit de VS, lijkt me ook logisch daar
dr Fauci eerder al meldde dat gevaccineerden nog lang besmettelijk
blijven en zich aan de Coronamaatregelen dienen te houden, terwijl de
voormaligfe minister van Volksgezodnheid, CDA plork fde Jonge vorig jaar
zelf toegaf dat gevaccineerden niet veilig zijn…… (zie de
afbeelding met deze zwaar disfunctionerende kwezel in het blok met Pfizer links hierboven)
‘Coronavirus, PCR test: leugens en onzin‘ (een artikel van Mario Ortiz Buijsse)
‘Met COVID-19 besmette zorgmedewerkers mogen ondanks symptomen gewoon doorwerken…….‘ (!!!!)
‘Coronavirus: alarmerend aantal dodelijke slachtoffers mRNA vaccins‘ (!!!!) (en zie ook de reactie onder dat bericht)
‘Coronacrisis: Australische militairen moeten de straten en huizen van o.a. moslims in Sydney controleren‘ zie wat betreft Australië en de Coronamaatregelen ook:
‘Frankrijk en Griekenland voeren vaccinatieplicht in voor verpleegkundigen en verzorgenden‘ (en zie de links in dat bericht!!)
‘COVID-vaccin slachtoffers openen website na censuur van media die imago van vaccinmakers belangrijker achten dan volksgezondheid‘ (!!!!) (en zie de links in dat bericht!!)
———————————————–
Over de betrouwbaarheid van de farmaceuten (zie daarvoor ook het eerste blok met links over Pfizer): ‘British Medical Journal eist onmiddellijke vrijgeving van alle gegevens over COVID-19 vaccins en onderzoek naar veiligheid‘
‘COVID-19 vaccin ontwikkeld in Texas: patentvrij voor de wereld‘ Terwijl de farmaceutische maffia (de grote farmaceuten) weigeren om hun recept gratis door te geven aan arme landen……
‘COVID vaccinmakers: hoe betrouwbaar zijn deze farmaceuten‘(!!!!)
‘COVID-19: internist, cardioloog en epidemioloog wijst op gevaar van vaccins‘
‘Coronavaccinatie ook als je weet dat de fabrikant ernstige bijwerkingen niet uitsluit?‘ (!!!!) en zie:
‘Coronavaccinatie
in vergelijking met het kopen van een huis (waar farmaceuten elke
verantwoordelijkheid voor bijwerkingen afwijzen!!)‘ (en zie de links in dat bericht)
‘Coronavirus:
Diederik Gommers (IC arts) zou het mooi vinden dat mensen worden
ingeënt met het AZ vaccin nadat ze tekenden voor het risico dat ze
lopen…..‘ (!!!!) (en zie de links in dat bericht!!) Een
bericht gebracht nadat bleek dat veel mensen slecht reageerden op het
AstraZeneca vaccin…. Eenzelfde eis overigens die de grote farmaceuten
aan de landen hebben gesteld waar hun vaccin werd verspreid, dus geen
gerechtelijke vervolging voor ernstige bijwerkingen op de korte,
middellange en lange termijn, kortom zelfs de makers vertrouwen hun
eigen vaccin niet……
‘COVID-vaccin slachtoffers openen website na censuur van media die imago van vaccinmakers belangrijker achten dan volksgezondheid‘ (en zie de links in dat bericht!!) (!!!!)
‘Coronabeslommeringen:
AstraZeneca vaccin onder vuur >> universiteit van Oxford start
opnieuw met angstzaaien voor het virus‘ (universiteit van Oxford heeft het vaccin gemaakt met AstraZeneca…….)
—————————————–
Wat betreft de betrouwbaarheid van de PCR test zie: PCR test in VS meer en meer onder vuur, ook de FDA en CDC willen ermee stoppen‘ en zie ook de volgende 5 links naar berichten over de PCR test en de waardeloze test op antilichamen:
‘Coronavirus:
PCR-test kan niet aantonen dat het virus aanwezig is in het lichaam,
beweringen dat dit wel zo is zijn gelogen‘ In dat bericht zie je een groot aantal links naar oudere artikelen
over het Coronavirus)
‘Uitvinder PCR test (algemeen gebruikt voor COVID-19): onbetrouwbaar als medische test‘ Hier een video over deze zaak met de uitvinder en de daarvoor met de Nobelprijs beloonde Kary Mullis:
‘Coronavirus, PCR test: leugens en onzin‘ Een artikel van Mario Ortiz Buijsse.
Ook de test op antilichamen klopt
niet, dit is een test voor mensen die het virus onder de leden hebben
gehad (!!!!):
‘COVID-19
testen op antilichamen van o.a. Roche, Quest en Abbott zijn
waardeloos‘
 Pfizer is under fire for secretive contracts with countries buying its Covid-19 vaccines. (Photo: Mario Tama/Getty Images)
Pfizer is under fire for secretive contracts with countries buying its Covid-19 vaccines. (Photo: Mario Tama/Getty Images)